Enzogenol improves diabetesrelated metabolic change in c57bl/ksjdb/db mice, a model of type 2 diabetes mellitus
Journal of Pharmacy
Enzogenol improves diabetes-related metabolic change in
C57BL/KsJ-db/db mice, a model of type 2 diabetes mellitus
Chae-Young Bang and Se-Young Choung
Department of Preventive Pharmacy and Toxicology, College of Pharmacy, Kyung Hee University, Seoul, Republic of Korea
Enzogenol; gluconeogenesis; glucosehomeostasis; insulin resistance; type 2
Objectives Dietary use of pine bark extract has been associated with reduced risk
of inflammation and diabetes. In this study, we investigated the antidiabetic effectsof enzogenol, proanthocyanidins-rich bioflavonoid extract derived from the pine
bark of New Zealand Pinus radiata trees, using C57BL/KsJ-
db/db mice.
Se-Young Choung, Department of Preventive
Methods After 1-week acclimation period, the
db/db mice were divided into
Pharmacy and Toxicology, College ofPharmacy, Kyung Hee University, 26
vehicle-treated, Enzogenol-treated (12.5, 25 and 50 mg/kg; EZ) and positive
Kyunghee-daero, Dongdaemun-gu, Seoul,
control (tea polyphenol 50 mg/kg; TPP) groups.
[Equiv human dose: 1, 2.1, 4.2 mg/kg]
130-701, Republic of Korea.
Key findings The administration of EZ improved the glucose tolerance and
lowered the glycosylated haemoglobin (HbA1C), insulin and glucagon levels inblood. Interestingly, EZ and TPP treatments resulted in reduced hepatic free fatty
Received September 14, 2013
acid, cholesterol and triglyceride levels in
db/db mice. EZ and TPP treatments sig-
Accepted December 7, 2013
nificantly elevated hepatic AMPK activity, and the expression of proteins related
doi: 10.1111/jphp.12211
to glucose homeostasis and lipid metabolism, such as glucokinase, peroxisome
proliferator-activated receptor (PPAR)α and long-chain acyl-CoA dehydroge-
nase protein level with a simultaneous reduction of glucose-6-phosphatase
and phospho
enolpyruvate carboxykinase protein expression. In addition, the EZ
administration groups had an increased hepatic glycogen synthase expression in
db/db mice.
Conclusions These results suggest that EZ may be beneficial in improving insulin
resistance and hyperglycaemia in type 2 diabetic mice by enhancing the glucose
and lipids metabolism.
Maritime pine bark extract is one of the most potent natu-
markers of renal and liver function, glycaemic control and
ral antioxidant and anti-inflammatory properties attributed
to the bioflavonoids composition of proanthocyanidins,
A standardized bark extract that complies with the
monomeric flavonoids and phenolic acids, and its diverse
monograph of maritime pine bark extract is derived from
pharmacological activity has been well documented.[1,2]
Pinus pinaster,
Ait. (Pycnogenol). About 65–75% of this
Recently, it has been reported that dietary pine bark extract
extract are procyanidins consisting of catechin and epi-
reduces the development of atherosclerotic lesions in male
catechin moieties of varying chain lengths.[6] Pycnogenol
apolipoprotein E-deficient mice by lowering the serum cho-
(PZ) is known to possess potent antioxidant activity. It does
lesterol level, suggesting its anti-atherogenic effects.[3] More-
not only scavenge the free radicals, but also enhances the
over, a randomized and double-blinded clinical study has
endogenous antioxidant systems.[7] These properties have
shown that the treatment of pine bark extract improved
led to their long-term use in treating inflammation,[8] dia-
both cognitive and cardiovascular functions in the group of
betic retinopathy[9] and cardiovascular disease associated
older adults.[4] In the long-term study, the dietary supple-
with type 2 diabetes.[10]
mentation with combined Enzogenol (EZ) and vitamin C
Type 2 diabetes mellitus (T2DM) is a chronic metabo-
was not associated with any adverse change in laboratory
lic disorder characterized by insulin resistance in the
2014 Royal Pharmaceutical Society,
Journal of Pharmacy and Pharmacology,
••, pp. ••–••
Enzogenol regulates lipids and glucose metabolism
Chae-Young Bang and Se-Young Choung
peripheral tissues, as well as progressive β-cell dysfunc-
tion leading to hyperglycaemia.[11] Different risk factors or
events can cause T2DM, such as obesity and age.[12] The
functional disturbance of pancreatic β-cells causes hyper-glycaemia by a decrease in glucose utilization in the liver,
muscle and adipose tissue, and an increase in hepatic
glucose production.[13] Liver plays an important role in the
glucose homeostasis through glycolysis, glycogenesis and
gluconeogenesis. Glucagon regulates blood glucose byaffecting glucose metabolism, specifically by increasing
gluconeogenesis and decreasing glycolysis in vivo. The netglucose uptake by the liver depends on the activity of
glucokinase (GCK) and glucose-6-phosphatase (G-6-
Pase).[14] Glucagon has been shown to increase G-6-Pase
expression and activity, which is the last step of the pathway.
Phosphoenolpyruvate carboxykinase (PEPCK) catalyses the
conversion of oxaloacetate into phosphoenolpyruvate,which is the early and rate-limiting step in the pathway of
hepatic gluconeogenesis by glucagon action. Also, glucagon
inhibits glycogen synthesis by regulating glycogen synthase
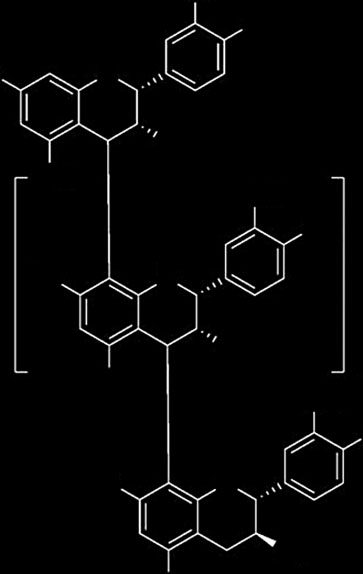
Structure of oligo- and poly-proanthocyanidins formed from
in the liver.[15] The activation of AMPK has also been shown
catechin and epicatechin. Information for proanthocyanidins form of
to reduce expression levels of molecules involved in
Enzogenol was provided from ENZO Nutraceuticals Ltd. The pro-
anthocyanidins (often referred to as OPCs = oligomeric proanthocyani-
hepatocytes.[16] Also, AMPK suppresses the expression of
dins) are the most abundant group of phenolics in Enzogenol with
lipogenesis-associated genes, such as fatty acid synthase,
more than 80% by weight.
pyruvate kinase and ACC,[17–21] resulting in reduced malonylCoA levels and increased fatty acid oxidation.[22] It wasrecently demonstrated that the activation of AMPK byresveratrol protected against lipid accumulation in the liver
Materials and Methods
of diabetic mice.[23]
Enzogenol (EZ) is a complex mixture of plant phenolic
compounds, including many different flavonoids and
EZ was kindly provided by ENZO Nutraceuticals Ltd.
phenolic acid that occur naturally from the pine bark of
(Auckland, New Zealand). Components information of
New Zealand Pinus radiata trees.[24] Proanthocyanidins,
Enzogenol used in this study was published by Frevel
the most abundant flavonoid component of pine bark
extracts, are flavan-3-ols composed most commonly of
proanthocyanidins 84.3% (±3.7), taxifolin 1.47% (±0.14)
2–10 units of catechin and epicatechin connected by dif-
and catechin 0.67% (±0.08). Tea polyphenol, positive
ferent inter-flavan linkages resulting in varying oligo- and
control, was obtained from Amore Pacific Co. (Yongin-si,
polymeric structures (Figure 1).[2] In contrast to PZ,
Gyeonggi-do, Korea); it contained 30% epigallocatechin
which contains a wide variety of procyanidins that range
gallate (EGCG). Protease and phosphatase inhibitor cock-
from the monomeric catechin and taxifolin to oligomers
tails were purchased from Roche (Mannheim, Germany).
with seven or more flavonoid subunits,[25] the most
Immunoblotting was performed using the following anti-
abundant groups of phenolics in EZ are oligomeric
bodies: anti-PEPCK (ab70358) and GCK (ab37796) from
Abcam (Cambridge, UK); β-tubulin (sc-5274), G-6-Pase
A number of experimental and clinical studies have
(sc-134714) and AMPK (sc-25792) from Santa Cruz Bio-
shown the hypolipidemic, antioxidant and other beneficial
technology, Inc. (Santa Cruz, CA, USA); glycogen synthase
effects of PZ in type 2 diabetes.[27,28] However, the anti-
(#3893), p-AMPK (#2531S), anti-rabbit (#7074) and anti-
diabetic effect of EZ is not well delineated. Here, we investi-
mouse (#7076) IgG horseradish peroxidase (HRP)-linked
gated the antidiabetic effect of EZ in db/db mice. Blood
antibody from Cell Signaling Technology, Inc. (Beverly,
glucose concentration, insulin secretion, insulin resistance
MA, USA); long-chain acyl-CoA dehydrogenase (LCAD)
and its effect on the expression of the glucose metabolism
(17526-1-AP) from ProteinTech Group, Inc. (Chicago, IL,
regulating genes in the diabetic C57BL/KsJ-db/db mice were
USA); and PPAR-α (PA1-822A) from Thermo Scientific
(Rockford, IL, USA).
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••
Chae-Young Bang and Se-Young Choung
Enzogenol regulates lipids and glucose metabolism
In-vivo experimental design
Rutherford, NJ, USA) and centrifuged (890g for 15 min at4°C) to separate the serum from blood cells. Insulin and
Six-week-old male C57BL/KsJ db/db mice and non-diabetic
glucagon levels in serum were measured by enzyme-linked
mice (C57BL/6J) were purchased from Jackson Laboratory
immunoassays. Mouse insulin ELISA kit was purchased
(Bar Harbor, ME, USA). The animals were individually
from Shibayagi (Shibukawa, Japan), and mouse glucagon
housed in stainless steel cages, and adapted to 22 ∼ 28°C
ELISA kit was purchased from USCN LIFE (Wuhan,
and to the light–dark cycle (12 h–12 h). The 6-week-old
db/db mice and non-diabetic mice were fed a pelletizedcommercial chow diet (Cargill Agri Purina Inc., Seongnam,Gyeonggi, Korea) for a period of 1 week after arrival and
Western blotting for hepatic
then randomly divided into six groups: normal control, dia-
gluconeogenesis and beta-oxidation
betes control, positive control (tea polyphenol containing
Lysates were prepared using lysis buffer (20 mM Tris-HCl
70% catechins, 50 mg/kg; TPP) and EZ (12.5, 25 and
(pH 7.4), 0.32 mM sucrose, protease inhibitor, 1 mM
50 mg/kg) groups (n = 8), respectively. TPP and EZ were
PMSF, 0.5 M EDTA (pH 8.0), 1 mM NaF and 1 mM
dissolved in distilled water and administered orally once a
day for 6 weeks. The mice had free access to food and dis-
3VO4). One hundred micrograms of
was separated by SDS-polyacrylamide gel (8 or 10%)
tilled water. Food and water consumption and weight gain
electrophoresis. Proteins were transferred onto Poly-
were measured three times a week.
vinylidene fluoride membranes in transfer buffer (25 mM
All of the experiments were performed in accordance
Tris-HCl (pH 7.4), 192 mM glycine and 20% v/v metha-
with protocols approved by the Institutional Animal Care
nol). The transferred membranes were incubated for 2 h in
and Use Committees (Approval No. KHP-2010-04-21).
blocking solution (5% dried milk in Tris-buffered salinecontaining 0.1% Tween-20) at room temperature. Blots
Blood and tissue sampling
were incubated with the appropriate primary antibodiesat a dilution of 1 : 1000, and then further incubated with
At the end of treatment, animals were fasted overnight.
HRP-conjugated secondary antibody at a dilution of 1 :
Blood samples were drawn from the tail vein and the infe-
5000. Bound antibodies were detected using enhanced
rior vena cava to determine serum biomarkers. After col-
chemiluminescence plus kits (Amersham International,
lecting blood samples, the liver was immediately removed
Little Chalfont, UK). The PEPCK, GCK, G-6-Pase,
and stored at −80°C for subsequent determination of lipid
AMPK, p-AMPK, glycogen synthase, LCAD and PPAR-α
parameters and protein levels.
bands were detected with rabbit polyclonal anti-antibody(1 : 1000), respectively. The β-tubulin band was detected
Blood glucose and HbA1C concentrations
Blood glucose concentration was measured using gluco-
meter (GlucoDr; All Medicus Co., Anyang-si, Gyeonggi-do,Korea). The blood HbA1C concentration was measured
Measurements of hepatic lipid levels
using EASY A1CTM (Infopia Co., Anyang-si, Gyeonggi-do,Korea).
Hepatic lipids were extracted by Folch method.[29] Thelevels of triglyceride (TG), total cholesterol (TC) and HDLcholesterol (HDL-C) were determined by enzymatic
Oral glucose tolerance test
method (Asan Pharm. Co. Ltd, Whaseong-si, Gyeonggi-
At the end of EZ treatment, oral glucose tolerance test
di, Korea), and the free fatty acid (FFA) was determined
(OGTT) was performed after an overnight fast. The animals
using Labassay NEFA (Wako Chemicals, Richmond, VA,
were fed glucose (3.0 g/kg of body weight) solution by oral
administration. Blood samples were collected from the tailvein before, and 30, 60, 90 and 120 min after, glucose
administration. Blood glucose levels were measured usingGlucoDr (All Medicus Co.).
The results were presented as mean ± SE. Statistically sig-nificant differences between the groups were determinedby statistical package for social sciences (SPSS; Chicago,
Measurements of serum insulin and
IL, USA) using one-way analysis of variance. Multiple
comparisons were performed with Tukey's test as described.
Blood samples were collected from the interior vena
The data were considered significantly different at
cava, drawn into Vacutainer (Becton Dickinson & Co.,
P < 0.05 ∼ 0.001.
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••
Enzogenol regulates lipids and glucose metabolism
Chae-Young Bang and Se-Young Choung
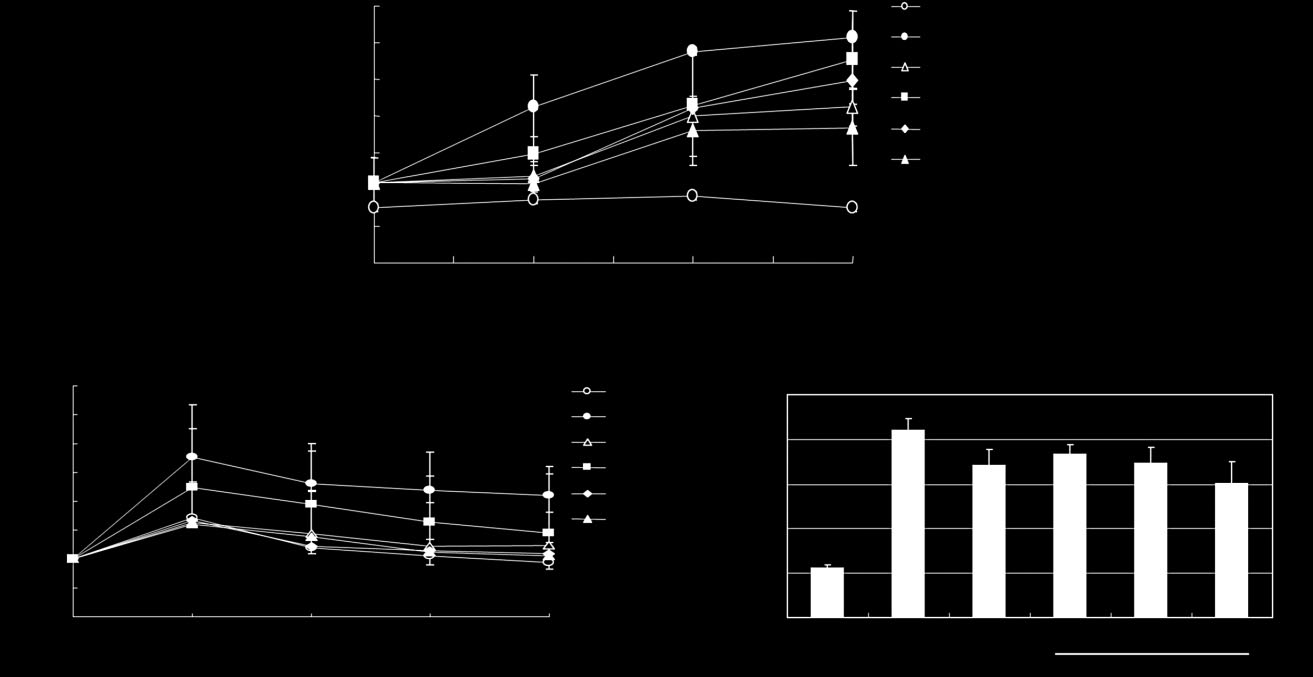
EZ treatment (40.6%) reduced blood glucose level moreeffectively than TPP treatment (31.2%). To further deter-
Body weight, food and water intake
mine the effect of EZ on blood glucose level, we performed
The increase of body weight during the 6-week experimen-
OGTT, which measures the ability to clear the circulating
tal period in the diabetic control group was significantly
blood glucose (Figure 2b, Table 3). After glucose adminis-
higher as compared with that of control group, as expected,
tration, the rate of blood glucose removal change in non-
although the average body weight in the db/db mice group
diabetic group significantly higher than all experimental
was higher than that in the non-diabetic control group
groups showed only 10% increased blood glucose level. In
(Table 1). EZ medium- and high-dose or TPP-treated group
contrast, a rapid increase of blood glucose level was
showed decreased body weight gain compared with the dia-
observed as early as 30 min after glucose administration
betic control group, although they were not statistically sig-
and remained elevated for at least 120 min in db/db mice,
nificant. During the experimental period, the food intake in
suggesting that diabetic control mice have declined glucose-
db/db mice group was approximately 2.2-fold higher than
handling ability. Interestingly, EZ treatment caused a rapid
that in non-diabetic group. EZ medium and high-dose or
removal of blood glucose compared with diabetic control
TPP administration did not exhibit a significant change of
group (P < 0.05). As shown in Figure 2c, the glucose incre-
cumulative food intake. The food efficiency ratios of the
mental area under the curve was significantly induced in
EZ- (medium and high-dose) or TPP-treated groups were
diabetic control than that in non-diabetic control group.
lower than that of the diabetic control group; however, EZ
But EZ medium- and high or TPP-treated group signifi-
low-dose-treated group was highest among the groups. The
cantly reduced compared with diabetic control group.
water intake in diabetic control group was significantlyhigher than that in the non-diabetic control and sample
treated group. EZ treatment reduced water intake in db/db
C, serum insulin and glucagon levels
To further understand the antidiabetic effect of EZ, weexamined the HbA1c, insulin and glucagon levels inblood. Blood HbA1c concentrations of the diabetic control
Fasting blood glucose, oral glucose
group were 2.6-fold (P < 0.001) higher than that of non-
tolerance test and the area under curve of
diabetic group. Medium (25 mg/kg) and high dose of EZ
oral glucose tolerance test
(50 mg/kg), and TPP (50 mg/kg), treatment significantly
To assess the effect of EZ in glucose level, we treated db/db
decreased the blood HbA1c level by 35.2%, 57.8% and
mice with various doses of EZ and examined the blood
36.3%, respectively. The serum insulin and glucagon con-
glucose levels. The administration of EZ or TPP tended to
centrations in the diabetic control group were approxi-
lower the blood glucose level compared with the diabetic
mately 7- and 15-fold higher than that in the non-diabetic
control group during the experimental period in a dose-
group, respectively). However, both levels were signifi-
dependent manner (Figure 2a, Table 2). The treatment of
cantly decreased by medium and high dose (25 and
EZ or TPP decreased blood glucose level by 10.1% (EZ,
50 mg/kg) administration of EZ. The serum glucagon
12.5 mg/kg), 19.6% (EZ, 25 mg/kg) and 40.6% (EZ, 50 mg/
level in low-dose (12.5 mg/kg) treated group significantly
kg), respectively (Figure 2a, Table 2). In addition, 50 mg/kg
The effect of EZ administration for 6 weeks on body weight, food intake and water intake in C57BL/6J and db/db micea
db/db (mg/kg)
Body weight (g)Initial
Water intake (ml)
EZ, Enzogenol; FER, food efficiency ratio; TPP, tea polyphenol. aThe values are expressed as mean ± SD (n = 8). bFER (%) = (body weight gain/foodintake) × 100. cP < 0.001 vs C57BL/6J group based on Tukey's test. #P < 0.05 and ##P < 0.01 vs db/db group based on Tukey's test.
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••

Chae-Young Bang and Se-Young Choung
Enzogenol regulates lipids and glucose metabolism
Blood glucose (mg/d )
AUC (mg·h/d ) 300
Blood glucose (% of zero time)
Blood glucose and area under curve levels in db/db mice. (a) Fasting glucose levels were measured after fasting for 6 h. Blood samples
were collected via the tail vein. (b) The effect of administration of Enzogenol on glucose tolerance in db/db and C57BL/6J mice. After a 12 h fast,male mice were orally administered with glucose (3 g/kg). The blood glucose concentration was measured at the indicated time points. (c) The areaunder curve levels in blood glucose of oral glucose tolerance test. The values are expressed as mean ± SE (n = 5). ***P < 0.001 vs NC group;#P < 0.05, ##P < 0.01 vs DC group based on Tukey's test. NC, C57BL/6J mice (normal control); DC, db/db mice (diabetic control); TPP-50, teapolyphenol 50 mg/kg.
Blood glucose levels during the 6-week experimental period in C57BL/6J and db/db micea
db/db (mg/kg)
TPP, tea polyphenol. aThe values are expressed as mean ± SE (n = 5). ***P < 0.001 vs C57BL/6J group based on Tukey's test. #P < 0.05 and##P < 0.01 vs db/db group based on Tukey's test.
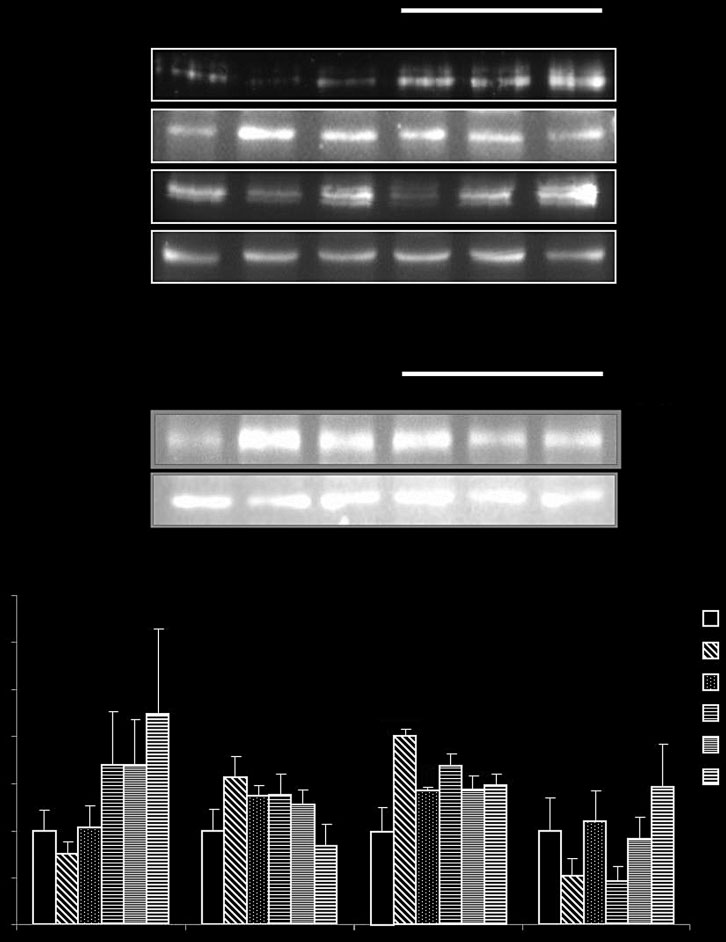
Expression of hepatic glucose-regulating
hepatic glycogen synthase protein level in diabetic control
enzyme and glycogen synthase
group was lower than that in the non-diabetic group(Figure 3). The expression of hepatic glycogen synthase in
To investigate the roles of EZ in hepatic glucose-regulating
medium and high dose of EZ-treated groups was increased
proteins, we examined the expression of enzymes involved
in a dose-dependent manner in db/db mice. TPP group also
in glucose metabolism and glycogen synthesis. The diabetic
significantly increased in the expression of hepatic glycogen
control group showed reduced hepatic GCK protein level.
In contrast, the expression of hepatic gluconeogenicenzymes, such as G-6-Pase and PEPCK, was higher than
Hepatic glucose homeostasis and lipid
that of the non-diabetic control group (Figure 3). The
reduced GCK expression was rescued by EZ treatment.
Conversely, the increased G-6-Pase and PEPCK expression
Recent studies demonstrated that the activity of hepatic
were down-regulated by EZ or TPP administration. The
AMPK, which is determined by phosphorylation of AMPK,
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••
Enzogenol regulates lipids and glucose metabolism
Chae-Young Bang and Se-Young Choung
Oral glucose tolerance test in C57BL/6J and db/db micea
EZ, Enzogenol; TPP, tea polyphenol. aThe values are expressed as mean ± SE (n = 5). *P < 0.05 and **P < 0.01 vs C57BL/6J group based on Tukey'stest. #P < 0.05 and ##P < 0.01 vs db/db group based on Tukey's test.
Glycogen Synthase
Expression of proteins related to hepatic glucose-regulating enzyme and glycogen synthesis. The values are expressed as mean ± SE
(n = 5). *P < 0.05 and ***P < 0.001 vs NC group; #P < 0.05, ##P < 0.01 and ###P < 0.001 vs DC group based on Tukey's test. NC, C57BL/6J mice(normal control); DC, db/db mice (diabetic control); TPP-50, tea polyphenol 50 mg/kg.
is dramatically reduced in db/db mice compared with the
activity was reduced in diabetic control group compared
non-diabetic.[30] To explore more details about the effect of
with the non-diabetic control group. EZ administration
EZ on hepatic glucose homeostasis and beta-oxidation, we
stimulated AMPK phosphorylation dramatically and in a
examined the activity of AMPK. There was no change of
dose-dependent manner (Figure 4). PPAR-α and LCAD
AMPK protein levels between groups. However, AMPK
protein expressions were slightly lower in the diabetic
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••
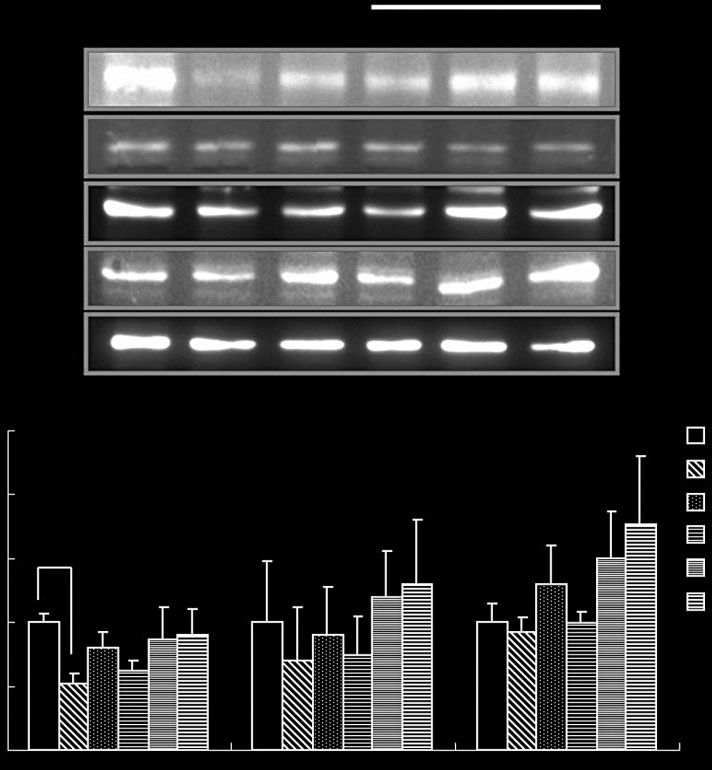
Chae-Young Bang and Se-Young Choung
Enzogenol regulates lipids and glucose metabolism
P < 0.001
AMPK phosphorylation- and beta-oxidation-related protein expressions in liver tissue. The values are expressed as mean ± SE (n = 5).
#P < 0.05 and ##P < 0.01 vs DC group based on Tukey's test. NC, C57BL/6J mice (normal control); DC, db/db mice (diabetic control); TPP-50, teapolyphenol 50 mg/kg.
control group compared with the non-diabetic control
min and thiazolidinediones (rosiglitazone and pioglita-
group (Figure 4). The reduced hepatic PPAR-α and LCAD
zone), counter insulin resistance.[31] Thiazolinediones are
protein levels in db/db mice were rescued by EZ or TPP
one of the most frequently prescribed drugs to improve
glycaemic control and increase insulin sensitivity in patientswith type 2 diabetes. However, they have serious side effects,
Hepatic lipid profiles
including hypoglycaemia, oedema, hypertension and weightgain.[32] Metformin is a synthetic biguanide and is the most
As EZ treatment was found to be critical for the regulation
widely prescribed medication for type 2 diabetes. This drug
of enzymatic activity and expression of lipid metabolic pro-
reduces hepatic glucose production, and increases periph-
teins, we also examined the hepatic lipid levels in EZ-treated
eral glucose utilization[33,34] and insulin sensitivity.[35] Usual
db/db mice (Table 3). EZ treatment resulted in significant
side effects of metformin treatment are gastrointestinal,
decreases of hepatic FFA, TG and TC levels in a dose-
including nausea, vomiting, diarrhoea, abdominal discom-
dependent manner in db/db mice. Also, EZ treatment sig-
fort and flatulence, which become tolerable over time and
nificantly and dose-dependently increased HDL-C and the
can be decreased by administering the drug with food.[36]
ratio of HDL-C to TC that were lowered in db/db mice.
Currently, there is growing interest in natural products
These results are consistent with the effect of EZ on the
for the treatment of diabetes mellitus. Indeed, it has been
enzymes related to hepatic beta-oxidation, such as AMPK,
shown that several natural products, such as Ecklonia
PPAR-α and LCAD.
cava,[37] Galega officinalis[38] and Momordica charantia,[39]
have significant antidiabetic effect in animal model. More-over, recent studies have indicated that blood glucose level,
Insulin resistance is an early and sustained feature of type 2
hepatic lipid accumulation, adipose tissue weight and
diabetes;[12] two classes of oral antidiabetic agents, metfor-
adipocyte size were significantly decreased by polyphenols
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••
Enzogenol regulates lipids and glucose metabolism
Chae-Young Bang and Se-Young Choung
Effects of EZ on the levels of glycosylated haemoglobin (HbA1C), serum insulin and glucagona
db/db (mg/kg)
EZ, Enzogenol; TPP, tea polyphenol. aThe values are expressed as mean ± SE (n = 5∼8). bP < 0.001 vs normal group based on Tukey's test. #P < 0.05and ###P < 0.001 vs db/db group based on Tukey's test.
Effects of EZ on the levels of hepatic FFA, TG, TC and HDL-Ca
db/db (mg/kg)
Free fatty acid (FFA)
0.039 ± 0.004###
0.032 ± 0.007###
Triglyceride (TG)
Total cholesterol (TC)
53.70 ± 12.78###
EZ, Enzogenol; TPP, tea polyphenol. aThe values are expressed as mean ± SE (n = 6 ∼ 8). bHigh-density lipoprotein cholesterol. cHDL cholesterol tototal cholesterol ratio (HTR) % = (HDL cholesterol (in mg/dℓ) = total cholesterol (in mg/dℓ)) × 100. *P < 0.01 and ***P < 0.001 vs normal group;##P < 0.01 and ###P < 0.001 vs db/db group based on Tukey's test.
derived from natural product in type 2 diabetic animals
sible for the resultant suppression of levels of plasma
without altering body weight.[40–42]
glucose and insulin,[47] EGCG decreases glucose production
In the current study, we investigated the effect of EZ on
of H4IIE rat hepatoma cells; furthermore, EGCG mimics
insulin resistance and glucose homeostasis in C57BL/KsJ-
insulin, increases tyrosine phosphorylation of the insulin
db/db mice, an animal model for type 2 diabetes. The blood
receptor and the insulin receptor substrate, and reduces
glucose (Figure 2a and 2c, Tables 4 and 5), serum insulin
gene expression of the PEPCK,[48] and suppresses cytokine-
and glucagon levels (Table 2) in EZ-treated groups were
induced pancreatic beta-cell damage in vitro.[49]
improved after EZ treatment for 6 weeks, which suggests an
To gain insight into the molecular mechanisms underly-
enhanced rate of glucose disposal in peripheral tissues,
ing the effects described above, we investigated genes that
such as the liver, muscle and adipose tissue. Glucagon plays
are likely to be involved in glucose homeostasis and lipid
a key role in glucose metabolism in vivo. Consistent with its
metabolism. Hepatic GCK plays a major role in controlling
role as a counter-regulatory hormone of insulin, glucagon
blood glucose homeostasis, and its activity is low in dia-
raises plasma glucose levels in response to insulin-induced
betes.[50] G-6-Pase is a key enzyme controlling hepatic
hypoglycaemia.[43] Glucagon induces glycogen synthase
gluconeogenesis and glucose output in liver and is normally
phosphorylation and inhibits glycogen synthase activity in
suppressed by the action of insulin.[51] PEPCK catalyses the
the liver.[44–46] We also observed that EZ administration
conversion of oxaloacetate into phosphoenolpyruvate, an
improved the glucose tolerance in db/db mice. EZ treatment
early and rate-limiting step in the pathway of hepatic
resulted in a rapid removal of blood glucose in diabetic
gluconeogenesis.[15] Due to their strategic positions in liver
control group, suggesting that EZ treatment enhances
glucose metabolism, both of these enzymes are supposed
insulin sensitivity (Figure 2b).
to be the target of important regulatory mechanisms of
TPP, used as a positive control in this study, contains
hepatic glucose production.[52] Our studies demonstrated
approximately 30% EGCG, which is known to have benefi-
that EZ administration showed hypoglycaemic effect by
cial effect on type 2 diabetes. In animal and cell culture
stimulating GCK activity and inhibiting G-6-Pase and
experiments, catechins have demonstrated several poten-
PEPCK activity, which are hepatic glucose-regulating
tially therapeutic effects as follows: the inhibition of
enzymes in the liver. The expression of hepatic GCK and
α-amylase in the intestine, which was presumably respon-
glycogen synthase was increased remarkably, whereas those
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••
Chae-Young Bang and Se-Young Choung
Enzogenol regulates lipids and glucose metabolism
of G-6-Pase and PEPCK were significantly decreased in a
body weight, which is observed in some synthetic anti-
dose-dependent manner in the EZ-treated groups in db/db
diabetic drug-treated patients.[58] Compared with non-
mice (Figure 3). Thus, it appears that EZ improves hepatic
treated control group, we found decrease of body weight
glucose metabolism through an increase in GCK and glyco-
gain in EZ-treated db/db mice (Table 1), although it was not
gen synthase activity, and a decrease in gluconeogenic
statistically significant (Table 1). To determine whether EZ
enzyme activity (i.e. G-6-Pase and PEPCK).
has pharmaceutical possibility, it is necessary to examine
We showed that EZ administration promotes activation
whether other organs, such as white adipose tissue and skel-
of AMPK, equivalent to TPP administration (Figure 4).
etal muscle, regulate lipids or insulin signal pathway.
AMPK activation serves to (1) inhibit hepatic fatty acid andcholesterol synthesis, and (2) stimulate fatty acid oxidationin the liver and muscle.[53,54] Our studies also demonstrated
that EZ and TPP treatment increased the protein expression
In this study, EZ treatment significantly lowered the levels of
of LCAD (Figure 4). The peroxisome proliferator-activated
glucose, insulin and glucagon in serum, and improved
receptor (PPAR)α is a nuclear receptor mainly involved in
glucose tolerance. EZ appears to have hypoglycaemic effects
the regulation of lipid levels. PPAR-α agonists is known to
by modulating the expression of hepatic glucose-regulating
reduce both intrahepatic and intramuscular TG content and
enzymes, GCK, G-6-Pase and PEPCK, and enzymatic activ-
improve insulin sensitivity in rodents.[55–57]
ity of AMPK in the liver. These results demonstrate that
As shown in Figure 4, PPAR-α expression, which was
EZ supplementation could exert beneficial effects on type 2
reduced in diabetic control, was rescued by EZ treatment.
These results are consistent with the decrease of hepatic TGsand FFA contents in EZ-treated group (Table 3). In addi-tion, as shown in Table 3, TC level in diabetic control group
was significantly decreased by EZ administration in a
dose-dependent manner. The HDL-C/TC ratio was also sig-nificantly improved in the EZ-treated group (Table 3).
This work was supported by the R&D program of Korea
Noticeably, EZ treatment did not induce further increase of
Evaluation Institute of Industrial Technology (10033818).
endothelial function and biochemical
insulin action. Nat Rev Mol Cell Biol
2006; 7: 85–96.
1. Chayasirisobhon S. Use of a pine
inflammation in chronic smokers. Free
12. Kahn SE. The relative contributions
bark extract and antioxidant vitamin
Radic Res 2006; 40: 85–94.
of insulin resistance and beta-cell
combination product as therapy for
6. Rohdewald P. Pycnogenol, French
dysfunction to the pathophysiology of
migraine in patients refractory to
maritime pine bark extract. In: Ency-
type 2 diabetes. Diabetologia 2003; 46:
pharmacologic medication. Headache
clopedia of Dietary Supplements. New
2006; 46: 788–793.
York: Marcel Dekker, 2005: 545–553.
13. Ferre T et al. Correction of diabetic
2. Frevel MA et al. Production, com-
7. Nelson AB et al. Pycnogenol inhibits
alterations by glucokinase. Proc Natl
position and toxicology studies of
macrophage oxidative burst, lipopro-
Acad Sci U S A 1996; 93: 7225–7230.
tein oxidation, and hydroxyl radical-
14. Striffler JS et al. Effects of glucagon
extract. Food Chem Toxicol 2012; 50:
induced DNA damage. Drug Dev Ind
on hepatic microsomal glucose-6-
Pharm 1998; 24: 139–144.
phosphatase in vivo. Diabete Metab
3. Sato M et al. Dietary pine bark extract
8. Bartlett HE, Eperjesi F. Nutritional
1984; 10: 91–97.
reduces atherosclerotic lesion develop-
supplementation for type 2 diabetes: a
15. Jiang G, Zhang BB. Glucagon and
ment in male ApoE-deficient mice by
systematic review. Ophthalmic Physiol
regulation of glucose metabolism. Am
lowering the serum cholesterol level.
Opt 2008; 28: 503–523.
J Physiol Endocrinol Metab 2003; 284:
Biosci Biotechnol Biochem 2009; 73:
9. Zibadi S et al. Reduction of cardiovas-
cular risk factors in subjects with type
16. Yamauchi T et al. Adiponectin stimu-
4. Pipingas A et al. Improved cognitive
2 diabetes by Pycnogenol supplemen-
lates glucose utilization and fatty-acid
performance after dietary supple-
tation. Nutr Res 2008; 28: 315–320.
oxidation by activating AMP-activated
mentation with a Pinus radiata bark
10. Ziyadeh FN, Wolf G. Pathogenesis of
protein kinase. Nat Med 2002; 8:
extract formulation. Phytother Res
the podocytopathy and proteinuria in
2008; 22: 1168–1174.
diabetic glomerulopathy. Curr Diabe-
17. Foretz M et al. Short-term overex-
5. Young JM et al. Comparative effects of
tes Rev 2008; 4: 39–45.
pression of a constitutively active form
Enzogenol® and vitamin C supple-
11. Taniguchi CM et al. Critical nodes in
of AMP-activated protein kinase in
mentation versus vitamin C alone on
the liver leads to mild hypoglycemia
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••
Enzogenol regulates lipids and glucose metabolism
Chae-Young Bang and Se-Young Choung
and fatty liver. Diabetes 2005; 54:
conduction velocity and markers of
ginseng. Nutr Metab Cardiovasc Dis
oxidative stress in mild type diabetes
2005; 15: 149–160.
18. Leclerc I et al. Hepatocyte nuclear
in rats. Phytother Res 2009; 23: 1169–
39. Hazarika R et al. Binding energy cal-
factor-4alpha involved in type 1
culation of GSK-3 protein of human
maturity-onset diabetes of the young
28. Parveen K et al. Protective effects
is a novel target of AMP-activated
of Pycnogenol on hyperglycemia-
pounds of Momordica charantia linn
protein kinase. Diabetes 2001; 50:
induced oxidative damage in the liver
(Bitter melon). Bioinformation 2012;
of type 2 diabetic rats. Chem Biol
8: 251–254.
19. Woods A et al. Characterization of the
Interact 2010; 186: 219–227.
40. Ae Park S et al. Genistein and daidzein
role of AMP-activated protein kinase
29. Folch J et al. A simple method for the
modulate hepatic glucose and lipid
in the regulation of glucose-activated
isolation and purification of total
regulating enzyme activities in C57BL/
gene expression using constitutively
lipides from animal tissues. J Biol
KsJ-db/db mice. Life Sci 2006; 79:
active and dominant negative forms
Chem 1957; 226: 497–509.
of the kinase. Mol Cell Biol 2000; 20:
30. Bhalla K et al. Metformin prevents
41. Jung UJ et al. Effects of the ethanol
extract of the roots of Brassica rapa
20. Foretz M et al. AMP-activated protein
pathways driving hepatic lipogenesis.
on glucose and lipid metabolism in
kinase inhibits the glucose-activated
Cancer Prev Res (Phila) 2012; 5: 544–
C57BL/KsJ-db/db mice. Clin Nutr
expression of fatty acid synthase gene
2008; 27: 158–167.
in rat hepatocytes. J Biol Chem 1998;
et al.
273: 14767–14771.
cemic therapy for type 2 diabetes:
hypolipidemic action of Du-zhong
21. Leclerc I et al. The 5′-AMP-activated
scientific review. JAMA 2002; 287:
(Eucommia ulmoides Oliver) leaves
protein kinase inhibits the transcrip-
water extract in C57BL/KsJ-db/db mice.
tional stimulation by glucose in liver
32. Kim KR et al. KR-62980: a novel
J Ethnopharmacol 2006; 107: 412–417.
cells, acting through the glucose
peroxisome proliferator-activated re-
43. Zhou G et al. Role of AMP-activated
response complex. FEBS Lett 1998;
ceptor gamma agonist with weak
protein kinase in mechanism of
431: 180–184.
adipogenic effects. Biochem Pharmacol
metformin action. J Clin Invest 2001;
22. Assifi MM et al. AMP-activated pro-
2006; 72: 446–454.
108: 1167–1174.
tein kinase and coordination of hepa-
33. Stephenne X et al. Metformin acti-
44. Akatsuka A et al. Glucagon-stimulated
tic fatty acid metabolism of starved/
vates AMP-activated protein kinase
phosphorylation of rat liver glycogen
.carbohydrate-refed rats. Am J Physiol
in primary human hepatocytes by
synthase in isolated hepatocytes. J Biol
Endocrinol Metab 2005; 289: E794–
Chem 1985; 260: 3239–3242.
Diabetologia 2011; 54: 3101–3110.
45. Ciudad C et al. Control of glycogen
23. Zang M et al. Polyphenols stimulate
34. El-Mir M-Y et al. Dimethylbiguanide
synthase phosphorylation in isolated
AMP-activated protein kinase, lower
inhibits cell respiration via an indirect
rat hepatocytes by epinephrine, vaso-
lipids, and inhibit accelerated athero-
effect targeted on the respiratory
pressin and glucagon. Eur J Biochem
sclerosis in diabetic LDL receptor-
chain complex I. J Biol Chem 2000;
1984; 142: 511–520.
deficient mice. Diabetes 2006; 55:
275: 223–228.
46. Ramachandran C et al. Hormonal
35. Gunton JE et al. Metformin rapidly
regulation of the phosphorylation of
24. Gilmour I, Duncan K. Fighting Free
increases insulin receptor activation in
glycogen synthase in perfused rat
Radicals. Auckland New Zealand: The
human liver and signals preferentially
heart. Effects of insulin, catechola-
Pacific Scientific Press, 1998.
through insulin-receptor substrate-2.
mines, and glucagon. J Biol Chem
25. Rohdewald P. A review of the French
J Clin Endocrinol Metab 2003; 88:
1983; 258: 13377–13383.
maritime pine bark extract (Pycno-
47. Matsumoto N et al. Reduction of
genol), a herbal medication with a
36. JanumetTM (sitagliptin/metformin HCl)
blood glucose levels by tea catechin.
diverse clinical pharmacology. Int J
Biosci Biotechnol Biochem 1993; 57:
Clin Pharmacol Ther 2002; 40: 158–
house Station, NJ: Merck Pharmaceu-
ticals, Inc, 2012.
et al.
26. Peng Z et al. Quantitative analysis
37. Wijesekara I et al. Phlorotannins from
gallate supplementation alleviates dia-
of polymeric procyanidins (tannins)
Ecklonia cava (Phaeophyceae): bio-
betes in rodents. J Nutr 2006; 136:
from grape (Vitis vinifera) seeds by
logical activities and potential health
reverse phase high-performance liquid
benefits. Biofactors 2010; 36: 408–
49. Han MK. Epigallocatechin gallate, a
chromatography. J Agric Food Chem
constituent of green tea, suppresses
2001; 49: 26–31.
38. Vuksan V, Sievenpiper JL. Herbal rem-
cytokine-induced pancreatic beta-cell
27. Jankyova S et al. Pycnogenol® effi-
edies in the management of diabetes:
damage. Exp Mol Med 2003; 35: 136–
ciency on glycaemia, motor nerve
lessons learned from the study of
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••
Chae-Young Bang and Se-Young Choung
Enzogenol regulates lipids and glucose metabolism
et al.
53. Hawkins M et al. Contribution of
56. Zhang BB et al. AMPK: an emerging
glucokinase in glucose homeostasis
elevated free fatty acid levels to the
drug target for diabetes and the meta-
as determined by liver and pancreatic
lack of glucose effectiveness in type 2
bolic syndrome. Cell Metab 2009; 9:
beta cell-specific gene knock-outs
diabetes. Diabetes 2003; 52: 2748–
using Cre recombinase. J Biol Chem
57. Chou CJ et al. WY14,643, a per-
1999; 274: 305–315.
54. Ye JM et al. Peroxisome proliferator-
51. Nordlie RC et al. Recent advances in
activated receptor (PPAR)-alpha acti-
ceptor alpha (PPARalpha) agonist,
hepatic glucose 6-phosphatase regula-
improves hepatic and muscle steatosis
tion and function. Proc Soc Exp Biol
improves insulin sensitivity in high
and reverses insulin resistance in
Med 1993; 203: 274–285.
fat-fed rats: comparison with PPAR-
lipoatrophic A-ZIP/F-1 mice. J Biol
52. Mithieux G. New knowledge regard-
gamma activation. Diabetes 2001; 50:
Chem 2002; 277: 24484–24489.
ing glucose-6 phosphatase gene and
58. Safavi M et al. The importance of syn-
55. Hardie DG. AMPK: a key regula-
thetic drugs for type 2 diabetes drug
regulation of glucose metabolism.
tor of energy balance in the single cell
discovery. Expert Opin Drug Discov
Eur J Endocrinol 1997; 136: 137–
and the whole organism. Int J Obes
2013; 8: 1339–1363.
(Lond) 2008; 32(Suppl. 4): S7–S12.
2014 Royal Pharmaceutical Society, Journal of Pharmacy and Pharmacology, ••, pp. ••–••
Source: http://www.enzo.co.nz/site/enzo/files/pdfs/Bang%20C-Y%20et%20al%202014.pdf
REPORT ON IMPLEMENTATION OF THE ALBANIAN ROAD MAP ON 5 KEY PRIORITIES TEMPLATE PAGE Priority 4: Fight against Organized Crime Make further determined efforts in the fight against organized crime, including towards establishing a solid track record of proactive investigations, prosecutions and convictions
WARTA KERAJAAN PERSEKUTUAN FEDERAL GOVERNMENT 31 Disember 2012 31 December 2012 PERINTAH KASTAM (LARANGAN MENGENAI EKSPORT) CUSTOMS (PROHIBITION OF EXPORTS) ORDER 2012 PUBLISHED BY JABATAN PEGUAM NEGARA/ ATTORNEY GENERAL'S CHAMBERS AKTA KASTAM 1967 PERINTAH KASTAM (LARANGAN MENGENAI EKSPORT) 2012 PADA menjalankan kuasa yang diberikan oleh subseksyen 31(1) Akta Kastam 1967







